

Introduction
Les Arthropodes sont les premiers animaux à avoir colonisé la terre ferme. Même si les premiers pas furent assez laborieux et que les essaies évolutifs furent nombreux, aujourd’hui, les Arthropodes forment un groupe cosmopolite, on les trouve dans des environnements naturels (déserts, forêts, abysses, montagnes…) ou d’origine anthropique (habitations, puits de pétroles, …). De plus il s’agit d’un groupe d’une étonnante diversité, environ 1 million d’espèces ont été découvertes et d’autres restent encore à découvrir.
Ils participent activement à la stabilité de l’écosystème en pollinisant les plantes, en décomposant la matière morte et en servant de base au réseau trophique.
Quels sont leurs particularités ? Comment ont-ils réussis à s’adapter à tous les milieux ?
I. Généralités
Les Arthropodes représentent le plus grand embranchement animal de la planète, ils regroupent 1,5 millions d’espèces. De plus beaucoups d’espèces restent encore à découvrir.
Etymologiquement le terme « Arthropode » vient d’arthron signifiant articulation et podos qui signifie pied, « Arthropode » veut dire « qui a des pieds articulés », ce groupe a été créé en 1845 par Siebold et Stannius. Sous ce nom sont groupés « les animaux de forme parfaitement symétrique, pourvus d'organes locomoteurs articulés et dont les masses centrales du système nerveux constituent un anneau ganglionnaire entourant l'œsophage et une chaîne ganglionnaire ventrale partant de cet anneau ».
Phylogénétiquement, les Arthropodes sont des :
- Métazoaires : Animaux pluricellulaires
- Eumétazoaires : Métazoaires possédant de vrais tissus
- Bilatériens : animaux possédant un axe de symétrie droite/gauche
- Protostomiens : pendant la vie embryonnaire, la bouche s’ouvre en premier
- Cuticulates : Animaux qui ont une cuticule à deux couches
- Ecdysozoaires : Animaux qui se développent par des mues successives

Cette classification n’a pas toujours été ainsi, en effet ce fut Latreille qui jugea bon de classer tous ces organismes ensemble, à partir de 1825, on classa tous les animaux instinctifs en un seul et même groupe, les Céphalidiens. Au sein de ce groupe il classa les Crustacés, les Arachnides et les Insectes en une race nommé les Condylopes, animaux articulés pourvus de pièces articulées. Ce terme correspond exactement à celui d’Arthropode créé vingt ans plus tard, mais il n’a pas eu la chance d’être retenu et de survivre.
Les Arthropodes sont à l’instar des Annélides, métamérisés, la présence de la cuticule va par ailleurs entrainer une croissance discontinue qui va contraindre l’animal à grandir via une suite de mue, les Arthropodes sont remarquables par 4 types de caractère :
- La présence d'une cuticule, aussi appelé exosquelette
- La tagmatisation
- Les appendices articulés
- L’adaptation à la vie terrestre
Du fait de l’originalité de ces caractères, l’origine des Arthropodes reste encore inconnue. Cependant leur ressemblance avec les Annélides est tout de même troublante, d’ailleurs le schéma d’un Annélide donne une idée de ce que peut être embryologiquement et approximativement un Arthropode. Ceci nous tente à rechercher dans le Précambrien des formes Annélides ayant une ressemblance avec ce groupe.
Les Euarthropodes sont divisés en 2 grands groupes, les Chélicériformes et les Mandibulates :
Les Chélicériformes forment un clade caractérisé par une division du corps en deux parties. La première étant le prosome constitué de l’acron et de six autres segments, chaque segment porte une paire d’appendices, la première de ces paires donnant les chélicères. La deuxième donnera les pédipalpes et les 4 autres donneront les 4 paires de pattes. La seconde partie étant l’opisthosome comportant jusqu’à 12 segments et le telson. Les Chélicériformes ont un système nerveux central ne comprenant qu’un protocerebron suivi du tricérébron autour de l’œsophage. Il n’existe pas de deutérocérébron comme chez les Mandibulates. Ces organismes sont divisés en deux autres clades :

Le sous-embranchement des Chélicérates qui constitue l’une des divisions majeures de l’embranchement des Arthropodes, ce taxon comprend les Arachnides et les Mérostomes (ici un regroupement de Limules). Ces animaux sont pour la plupart des prédateurs.
Les chélicérates, comme tous les chélicériformes ont un corps divisé en 2 parties. Le prosoma antérieur encore appelé céphalothorax composé de 8 segments auxquels s’ajoute une pièce supplémentaire antérieur, l’acron et l’opisthosoma postérieur (ou abdomen) constitué de 12 segments et d’une pièce supplémentaire, le telson.
Chez les chélicérates, les segments ne sont cependant pas visibles de l’extérieur à l’exception des Scorpionides au niveau de leur abdomen

Le second sous embranchement regroupe ce qu’on nomme les Pantopodes (ici Nymphon grossipes) ou Pycnogonides (les Pantopodes étant le seul ordre restant des Pycnogonides), ce sont des animaux de taille souvent inférieure au centimètre, leur corps est plat et segmenté, doté de quatre paires de pattes, ils sont munis d’une trompe parfois complétée par une paire de chélicères. Le caractère spécifique à ces Pycnogonides est la disparition partielle voire complète de l’opisthosome, du fait de cette absence, les organes normalement présents dans l’abdomen se retrouvent dans les pattes. D’ailleurs chez la femelle, il y a une paire de pattes utilisée principalement pour transporter les œufs jusqu’à leur éclosion qu’on appelle les ovigères.
Il existe un second groupe d’Euarthropodes, les Mandibulates, encore appelés Antennates dans certains ouvrages. Ces Mandibulates regroupent les Arthropodes dont la tête porte des appendices caractéristiques souvent très chitinisés et très durs, les mandibules. La monophylie de ce taxon s’explique aussi par l’étude moléculaire des ARNs ribosomiques (ARN 18S en particulier). Les Mandibulates sont divisés en 2 sous embranchements, les Myriapodes et les Pancrustacés. Ils étaient classiquement divisés en Uniramés (Insectes et Myriapodes) et Biramés (Crustacés). Cependant les similitudes entre les Myriapodes et les Insectes ont été expliquées comme des convergences liées à l’adaptation au milieu terrestre. Les insectes sont en effet plus proches des Crustacés.

Couramment appelés mille-pattes, les Myriapodes (ici une scolopendre mange un Lycodon laoensis) regroupent les Arthropodes composés d’une tête suivis de nombreux anneaux semblables, portant chacun une ou deux paires de pattes. Les Myriapodes sont tous des animaux terrestres, on peut cependant les trouver grimpant aux arbres soit pour se réchauffer soit pour se nourrir, ou encore pour chasser. Les Myriapodes forment un clade à la monophylie discutée. En effet les 4 classes qui le composent pourraient ne pas être apparentées.

Les Pancrustacés représentent un taxon récent issu de la phylogénie, il s’agit d’un taxon assez large qui regroupe plusieurs clades qui paraissent polyphylétiques, les Crustacés et les Hexapodes. Ce sous embranchement inclut donc les Crustacés au sens strict, c'est-à-dire les Rémipèdes, les Céphalocarides, les Maxilopodes, les Branchiopodes et les Malacostracés, les Pancrustacés regroupent aussi les anciens Hexapodes, c'est-à-dire, les Insectes, les Collemboles, les Protoures et les Diploures. La place des anciens Hexapodes n’est pas fermement établie à ce jour cependant toutes les études phylogénétiques tendent à les classer à proximité des Malacostracés et ou des Branchiopodes.
Le terme Hexapodes est toujours utilisé pour présenter les 4 classes précédemment citées, cependant il reste Phylogénétiquement incorrect.
Caprella septentrionalis (© Clione.ru) est une espèce d’Amphipode (crevette) vivant sur les algues, son corps parfaitement homochromique lui permet de se dissimuler dans les algues rouges où elle vit. Comme tous les membres de la famille des Caprellidae, elle est carnivore, elle chasse à l’affut telle une mante religieuse grâce à leurs appendices antérieurs transformés en crochets

Récemment les Protoures et les Collemboles ont été regroupés dans un taxon appelé Ellipoures, les Diploures quant à eux ont été scindés en 2 autres classes, les Campodéoïdes et les Japygoïdes. Il en est de même pour la classe des Myriapodes, des études récentes montrent qu’il n’existe aucune parenté entre les différents ordres.
II. Caractères généraux, Métamérie et coelome
A. Notions d’embryologie : réalisation de la métamérie
Le corps des Arthropodes est constitué d’éléments semblables qu’on nomme métamères, ceux-ci sont reliés les uns aux autres par de souples membranes. Ces métamères se forment au cours du développement embryonnaire de l’animal.
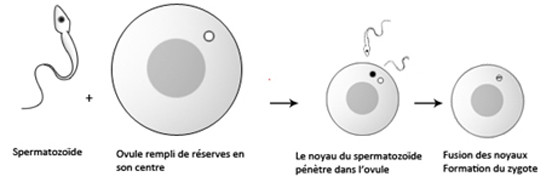
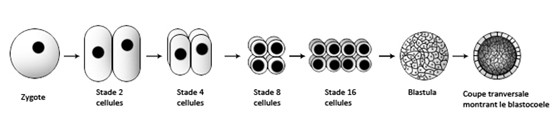
Lors de la fécondation, le spermatozoïde et l’ovule fusionnent conduisant à la formation d’une cellule œuf qu’on nomme le zygote, cette cellule se divise plusieurs fois pour devenir un embryon. Cet amas de cellules appelé blastula entoure une cavité qu’on nomme le blastocoele.
Pendant la gastrulation, les cellules de la blastula migrent dans le blastocoele de façon à s’organiser en 3 feuillets embryonnaires, l’ectoderme à l’extérieur, l’endoderme à l’intérieur et le mésoderme entre les deux autres feuillets.
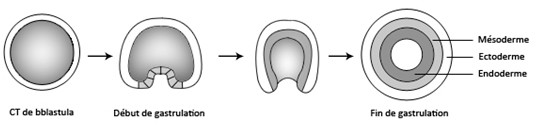
Le mésoderme se creuse d’une cavité, qu’on nomme le cœlome, celui-ci se scinde en plusieurs petites cavités.

Les métamères correspondent à la répétition tout le long du corps (de la tête à l’anus) de la même organisation cellulaire autour d’une paire de cavités cœlomiques.
Une fois les métamères formés, les parois des vésicules cœlomiques métamérisées vont se dissocier très rapidement. Ces vésicules vont alors fusionnées avec le blastocoele embryonnaire, une nouvelle cavité va donc se former, l’hémocoele, dans cet hémocoele circule un liquide qu’on nomme hémolymphe. Chez l’adulte il n’y aura plus grand-chose qui sera originaire du cœlome embryonnaire, le cœlome est en effet vestigial, il formera la cavité des glandes génitales avec une portion initiale, ou saccule, de certains organes excréteurs segmentaires comme le rein céphalique des anciens Thysanoures, la glande antérieure (ou mandibulaire) des Crustacés ou encore la glande coxale des Arachnides.
En se dégradant, les parois des vésicules cœlomiques vont devenir des muscles, ces muscles ont une organisation particulière chez les Arthropodes. En effet il n’y a plus de muscle circulaire, les Arthropodes ne se déplacent plus par les mouvements du corps mais par les mouvements de leurs pattes articulées.
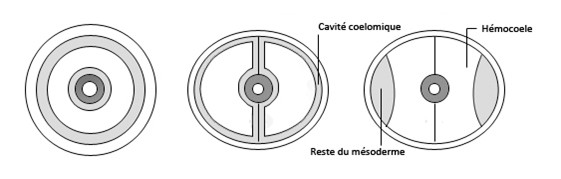
Du fait de cette fusion des vésicules cœlomiques avec le blastocoele, la métamérie chez les Arthropodes est difficile a observer à l’intérieur du corps, cependant elle reste plus ou moins visible à l’extérieur. Cette métamérie est encore altérée par la tagmatisation du corps.
B. Les métamères
Le corps des Arthropodes résulte donc de la juxtaposition longitudinale de métamères. La disposition régulière de ces unités structurales et fonctionnelles se traduit par une répartition périodique des organes ou fragments d’organes. Chaque métamère est encadré par plusieurs plaques squelettiques, une pièce dorsale, le tergite, une pièce ventrale, le sternite et deux pièces latérales souvent fusionnées, les pleurites qui sont dorsales et les épimérites qui sont ventrales.
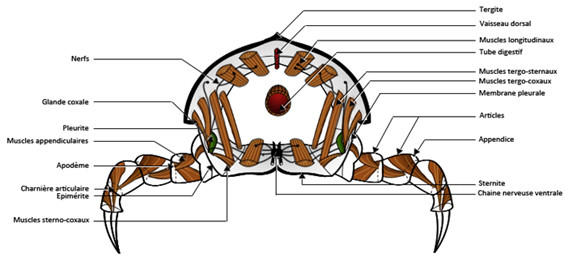
Ces différentes pièces sont réunies par des zones articulaires souples et minces qui sont également présentes entre les métamères. Le métamère est traversé par la chaine nerveuse ventrale, le tube digestif et le vaisseau dorsal. Il renferme une paire de ganglions, deux faisceaux musculaires dorsaux et deux faisceaux musculaires ventraux. Ces muscles sont insérés sur des replis de la cuticule, les apodèmes. Ces apodèmes marquent les limites antérieures et postérieures des métamères.
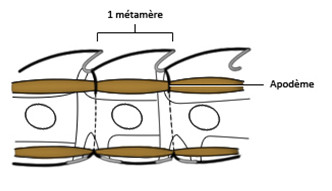
Chaque métamère porte deux appendices composés de plusieurs articles, ces appendices sont insérés entre les pleurites et les épimérites. Chaque métamère est donc un ensemble complexe comprenant :
- Un segment ectodermique limité par des sillons intersegmentaires
- Un segment vasculaire : le vaisseau dorsal
- Un segment digestif
- Un segment nerveux appelé également le neuromère
- Un segment musculaires (muscles ventraux et dorsaux et muscles des pattes) appelé aussi le myomère
- Un segment excréteur appelé aussi néphromère
- Une paire d’appendices appelée aussi segment appendiculaire
Tous ces éléments résultent d’une intégration anatomique et physiologique au sein du métamère. Tous les composants sont liés les uns aux autres.
Chez les Arthropodes, on distingue l’acron qui est l’équivalent du stomodeum des Annélides, le tronc ou soma composé de la juxtaposition des métamères vrais et le telson, équivalent au pygidium des Annélides. L’acron et le telson ne sont pas des métamères. L’acron n’est pas forcément visible, il renferme le cerveau primitif, qui est dorsal, le protocerebron encore appelé archicérébron. Il ne s’agit pas d’un métamère car il ne contient pas de sac cœlomique. La bouche s’ouvre en arrière de l’acron, au niveau du premier métamère vrai. Le telson est toujours bien visible, il est dépourvu de ganglions nerveux et de vésicules cœlomiques, il porte l’anus.
C. Les altérations de la métamérie. La tagmatisation
Contrairement aux Annélides, les divers métamères des Arthropodes sont dissemblables, on parle de métamérie hétéronome. Ils s’ordonnent de manière à former des régions morphologiquement distinctes. Cette spécialisation est accompagnée de la fusion des métamères dont les appendices collaborent à la réalisation de la même fonction (locomotion, mastication, reproduction, perception sensorielle).
Les métamères et les appendices associés qui constituent le soma vont fusionner pour former des tagmes, ces tagmes vont être morphologiquement différents et vont être spécialisés dans des fonctions. La tête est la zone des sens, elle sert à la perception sensorielle et à la prise alimentaire, le thorax sert à la locomotion et l’abdomen sert à la reproduction.
En fonction des différentes classes d’Arthropodes, les tagmes portent des noms différents.
1. Chez les Arachnides
L’Arachnide a un corps qui est divisé en deux régions distinctes : l’une antérieure, le prosoma ou céphalothorax, l’autre postérieure, l’opisthosoma ou abdomen.
Le prosoma porte les yeux, toujours simples, et six paires d’appendices chez l’adulte. La première paire représente les chélicères qui sont en position antérieure. Les chélicères sont suivies de la deuxième paire d’appendices, les pédipalpes (ou pattes mâchoires). Les quatres autres paires représentent les pattes à fonction ambulatoire. Le prosoma compte au minimum six métamères car il y a six paires d’appendices.
L’opisthosoma ne porte aucun appendice. Cependant il peut porter très rarement des formations spéciales d’origine appendiculaire comme des peignes ou des filières. Le nombre de métamères abdominaux varie de douze à quinze, selon les ordres.
Chez les scorpions et selon les auteurs, l’opisthosoma peut être divisé en deux parties, le mésosoma (ou pré-abdomen) qui fait suite au prosoma et qui compte 7 segments et le métasoma (surnommé queue) composé de 5 segments (dont le telson qui se finit en un aiguillon parfois venimeux)

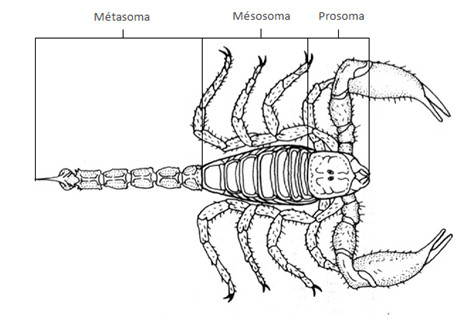
2. Chez les Myriapodes
Les Myriapodes sont des Arthropodes qui sont tous terrestres, ils se distinguent des autres Arthropodes par le fait que les métamères sont regroupés deux à deux de manière plus ou moins apparente, on parle de diplopodie. Chaque groupement est appelé diplosegment, il porte dans le cas le plus simple deux paires de pattes. Chez les Chilopodes, un ordre de Myriapode, les diplosegments ne portent qu’une seule paire d’appendices. Le corps des Myriapodes est divisé en deux parties, la tête et le tronc.

3. Chez les Crustacés
L’organisation des Crustacés est très proche de celle du plan d’organisation d’un Arthropode primitif, la tête ou céphalon, incluant au moins cinq segments d’origine somatique, le thorax, ou péréion, dont un certain nombre variable de segment peuvent être soudés à la tête, on parle de céphalothorax, la troisième partie du corps est l’abdomen ou pléon. Les appendices associés au péréion sont appelés péréiopodes, ceux associés au pléon sont appelés pléopodes. Le dernier segment est le telson, il forme souvent une nageoire caudale avec les deux derniers pléopodes, nommés uropode.
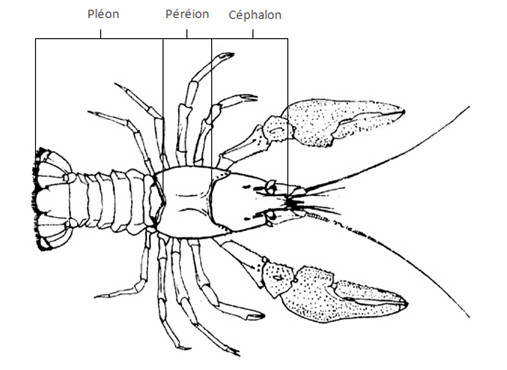
4. Chez les Insectes
Le corps des Insectes est quant à lui séparé en 3 parties :
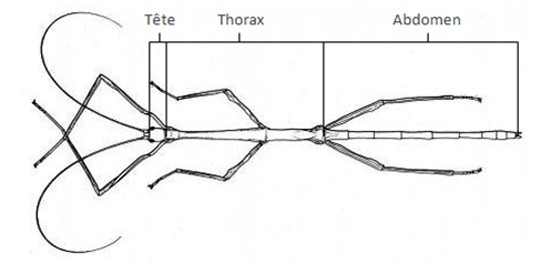
- La tête est une partie assez homogène où ne sont plus reconnaissables les six segments originels, l’acron et 5
métamères.
- Le thorax comprend le prothorax, le mésothorax et le métathorax. Chacun de ces trois segments porte une paire de pattes ambulatoires, les deux derniers, pouvant porter une paire d'ailes, constituent, chez les Ptérygotes, le ptérothorax.
- L'abdomen comprend originellement une douzaine de segments. Par suite de fusions ou de télescopages, il en comporte souvent un nombre visible bien moindre. En général, on peut admettre que l'abdomen se termine, par l'anus, sur le onzième segment, quant au processus génital, il est porté par le 9ème.
D. La céphalisation
La tête est retrouvée chez tous les Arthropodes, ce tagme résulte de la fusion de l’acron et d’un nombre variable de métamères selon la classe d’Arthropodes à laquelle on s’intéresse. Chaque paire d’appendices que portait chaque métamère composant la tête va perdre sa fonction ambulatoire originelle, ils vont se différencier en organes sensoriels, masticateurs ou préhenseurs (qui sert à la saisie).
La céphalisation ne laisse aucune trace de la fusion des métamères. En effet, la tête forme une capsule céphalique rigide qui présente des sutures plus ou moins apparentes qui sont, soit des traces externes d’apodèmes (renfoncement interne de l’exosquelette qui permet l’insertion des muscles), soit des lignes d’exuviations, ce sont des zones non résistantes où la cuticule de l’animal est très fine, c’est là que l’exosquelette va se fissurer lors de la mue. Ces sutures ne représentent aucunement la fusion des métamères, le dénombrement de ces métamères n’est possible que par une étude embryologique approfondie.
La céphalisation s’accompagne aussi de la fusion des ganglions nerveux appartenant à chaque métamère de la tête, il en résulte une structure nerveuse qui sera dorsale par rapport au tube digestif, cette structure est nommée cerveau, il innerve les organes sensoriels. Chez tous les Arthropodes actuels connus, le cerveau comporte trois neuromères (un neuromère est la partie nerveuse d’un métamère).
III. Originalité des arthropodes
A. La Paroi du corps
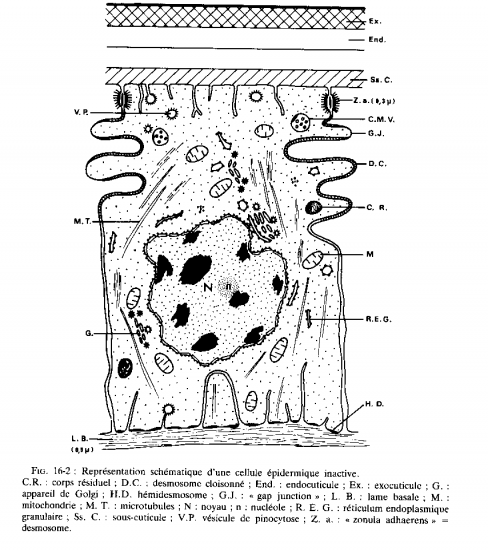
1. L’épithélium
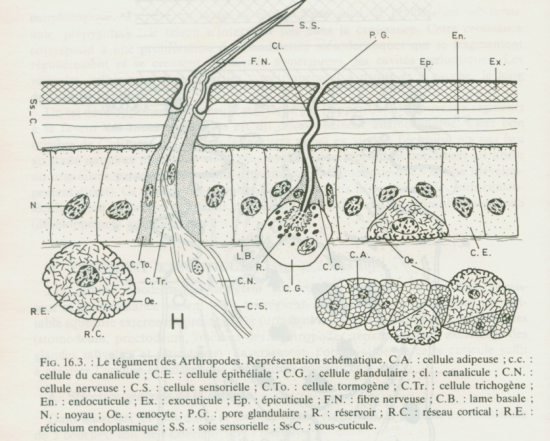
L’épithélium qu’on appelle aussi épiderme ou encore hypoderme selon l’ouvrage est constitué d’une seule couche de cellules cubiques, on parle d’épithélium cubique unistratifié, cet épiderme repose sur une lame basale percée de pores permettant le passage de plus grosses protéines venant de l’hémolymphe. La lame basale est un assemblage de protéines et de glycoprotéines (protéines associées à des sucres) extracellulaires, elle forme une séparation avec l’hémocoele sous jacent.
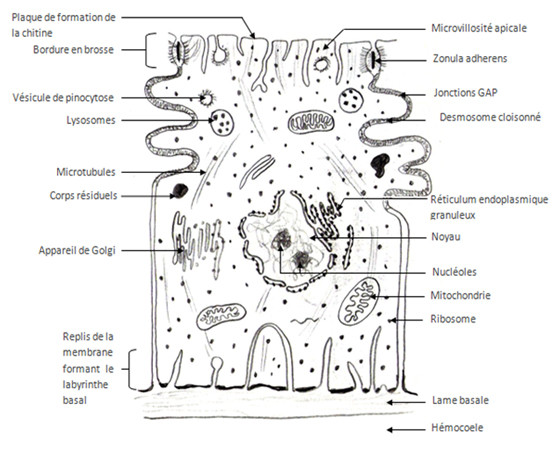
Zonula adherens, jonctions GAP et desmosomes cloisonnés sont impliqués dans la jonction cellulaire, ceci permet au tissu d’être résistant et imperméable à différents agents. Les hémi-desmosomes sont impliqués dans la jonction cellule-lame basale, ces points d’attache permettent aux cellules d’adhérer à la lame basale et de former un tissu très bien structuré.
Le noyau cellulaire contient l’ADN, l’ADN est coupé en gènes, chaque gène sera transcrit en ARNm (ARN messager) qui est en quelque sorte une copie du gène capable de sortir du noyau. Les nucléoles sont des zones de transcriptions intenses. Une fois formés, les ARNm sont traduits en protéines au niveau des ribosomes et au niveau du réticulum endoplasmique granuleux. Les protéines produites peuvent enfin être glycosylées (rajout de molécules de sucre) dans l’appareil de Golgi, elles seront ensuite utilisées à plusieurs fins (élaboration de structures cellulaires, hormones, etc.…)
Les mitochondries sont les usines à énergie de la cellule, c’est là que les nutriments avalés par l’animal seront transformés en énergie chimique utilisable pour la cellule.
La membrane cellulaire peut s’invaginer dans la cellule en emportant avec elles des éléments du milieu, ceci conduit à la formation de vésicules de pinocytose, concrètement, ces vésicules servent à « sonder » le milieu, c'est-à-dire qu’elles se tiennent informées des conditions extérieures. Ces vésicules peuvent fusionner avec des structures servant à la digestion intracellulaire, les lysosomes, on parle alors de phagolysosomes. En dégradant les molécules de la vésicule de pinocytose le lysosome peut se remplir de déchets non dégradables, on parle alors de corps résiduels.
Les cellules épithéliales sont les plus nombreuses, elles portent des microvillosités qui sont réduites et tronquées, elles portent aussi des plaques formant des épaississements du cytoplasme où a lieu la dernière étape de la synthèse de la chitine. Au niveau apical de ces cellules, on a des canaux qui vont se ramifier en canaux ciriers (sécrétant de la cire) certains canaux vont arriver jusqu’au coté apical de l’épicuticule et déverser leurs substances.
On trouve un deuxième type de cellules, les glandes épidermiques, prolongées aussi par un canal qui se finit au dessus de la cuticule.

Il y a un autre type de cellules qu’on appelle les oenocytes, elles fabriquent une substance cireuse qu’elles transfèrent aux cellules épidermiques, ces cellules se trouvent entre l’épiderme et le tissu adipeux sous jacent.
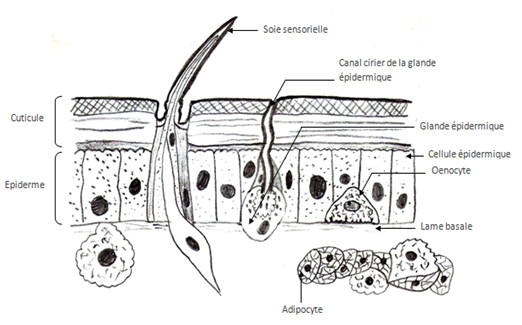
L’épiderme participe donc à la confection des soies, des épines présentes chez beaucoup d’Arthropodes (phasme à tiares, pinces de langoustine, etc.) ou encore d’écailles comme sur les ailes des Papillons. La couleur de ces animaux peut être due à plusieurs choses :
- Des pigments qui sont stockés dans l’épiderme ou dans les différentes couches de la cuticule.
- La structure physique de la cuticule, la manière dont la cuticule est agencée va faire que la lumière passera plus ou moins bien, la structure absorbant plus ou moins de couleur et nous renverra la couleur de l’animal. Les écailles des ailes des Papillons du genre Morpho d’une couleur bleue par exemple.
- Une combinaison de pigment et de structure physique.
2. La cuticule
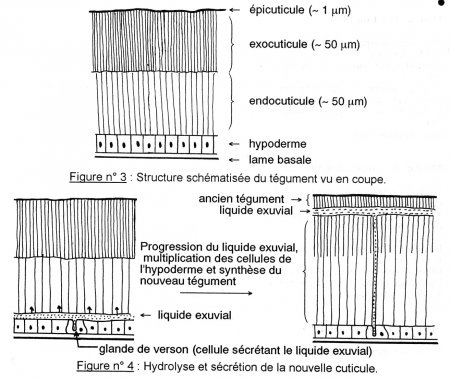
L’épiderme fabrique une substance répartie en 2 couches :
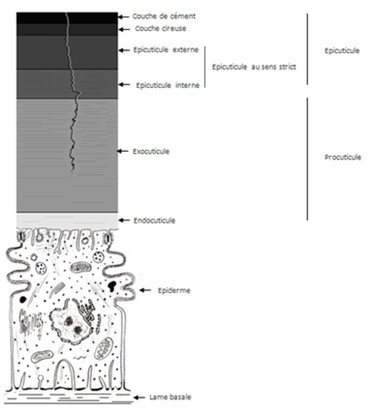
- L’épicuticule : elle se trouve à la surface de la cuticule, elle est en contact avec le milieu extérieur, mesure de 2 à 3 µm et est constituée de 3 sous couches, elle sert à imperméabiliser l’animal et à limiter les pertes d’eau par évaporation.
La couche de cément : cette couche est parfois absente, elle est constituée de lipoprotéines tannées produites par des glandes épidermiques dont le canal excréteur traverse toute l’épaisseur de la cuticule. Cette couche à une épaisseur d’1 µm d’épaisseur environ.
La couche cireuse appelée aussi couche moyenne de cires : faite de paraffine, d’acide gras et d’alcools à longues chaines, la cire produite par les oenocytes emprunte ensuite les canaux poraires (ou canalicules des pores) qui sont des digitations issus des cellules épidermiques et traversant toute l’épaisseur de la procuticule, ces canaux passent dans l’épicuticule et portent le nom de canaux ciriers. Cette couche mesure 0,5 à 1 µm d’épaisseur et permet de limiter les pertes d’eau, les Crustacés n’en possède pas car cette couche leur est inutile
Ces deux dernières couches sont qualifiées de couches postexuviales, elles ne seront sécrétées que quand l’animal changera de cuticule. Par opposition à la dernière couche que nous allons décrire.
L’épicuticule au sens strict : elle fait 0,5 µm d’épaisseur, et sert à la fixation des muscles et dans la transmission des signaux chimiques (aussi appelés phéromones). Cette couche est faite de lipoprotéines tannées qu’on appelle cuticuline, il s’agit d’une couche prééxuviale que l’on peut subdiviser en deux parties:
L’épicuticule externe qui mesure 20 nm d’épaisseur qu’on nomme encore cuticuline. En période d’intermue, elle a une structure trilaminaire
L’épicuticule interne mesurant de 0,2 à 0,5 µm d’épaisseur. Cette sous couche est riche en protéines, en composés aromatiques et contient des stérols.
- La Procuticule : Ce sont des couches qui contiennent de la chitine. La chitine est une substance synthétisée à partir du tréhalose, dimère de glucose regroupé par des liaisons α, α. 1-1, la chitine est un dimère de glucosamine qui polymérise dans l'hémolymphe en formant des filaments sur lesquels vont se greffer des protéines ou d'autres molécules. La chitine a un rôle protecteur, car lorsqu’elle est associée à du carbonate de calcium, elle devient très rigide et forme l’exosquelette des Crustacés et la coquille d’autres animaux. Cependant, ce n'est pas la chitine qui est le principal élément rigide de la cuticule, mais une protéine très particulière appelée la sclérotine qui la compose à 80%.
L’exocuticule : est sécrétée avant la mue, il s’agit donc d’une couche prééxuviale, elle est dure, rigide et colorée. En effet, elle présente des stratifications de couleur ambrée. Elle est insoluble dans l’eau, la chitine est associée à des protéines tannées qu’on nomme sclérotine, cette protéine rend la couche rigide colorée. La mélanine y est également présente pour donner la coloration brunâtre-noirâtre aux Arthropodes. Cette couche représente 1/12 à la moitié de l’épaisseur totale de la cuticule.
L’endocuticule est une couche contenant encore plus de chitine, ce qui la rend plus souple. De plus, elle est associée aux arthropodines qui sont aussi des protéines souples, cette association rend cette couche souple.
L’exocuticule et l’endocuticule sont percées par des canalicules de diamètre assez variable, les canalicules des pores aussi appelés canalicules poraires. Ces deux couches peuvent être séparées par une mésocuticule.
La procuticule est fréquemment séparée de l’épiderme par une couche assez anarchique composée de mucopolysaccharides acides, la sous cuticule. On ne connait pas encore le rôle de cette sous cuticule mais on suppose qu’il s’agit certainement d’une couche de « lubrifiant » qui sert aussi à l’osmorégulation des différents milieux. L’osmorégulation consiste à l’équilibre ionique entre les différents milieux.
3. Composition chimique
a. La chitine
La chitine est un polysaccharide azoté et acétylé qui représente environ 50% du poids sec de la cuticule des Arthropodes, c’est un composé qui se polymérise très facilement et qui porte le nom de poly-acétyl-D-glucosamine. Chimiquement, il s’agit d’un C8H13O5N répété n fois. L’hydrolyse de ce composé fournit du glucosamine.
Tout d’abord, une fonction acétyle s’associe au niveau de l’azote porté par le carbone 2 du glucosamine, cette association fournit de l’acétyl-glucosamine, il s’agit de l’élément unitaire conduisant à la fabrication de la chitine. Dans un second temps, deux de ces molécules s’assemblent par une liaison osidique pour former du tréhalose, ou chitobiose.
Au niveau de l’hémolymphe, la chitine se polymérise, les polymères s’assemblent parallèlement entre eux par des ponts latéraux en formant des micelles allongées à disposition pseudo-cristalline.
La chitine est une substance souple et perméable à l’eau du fait de sa nature sucrée. Il n’existe donc aucun rapport entre la richesse en chitine et la dureté du tégument. La disposition de la chitine fait qu’elle s’assemble à d’autres molécules, ce sont ces molécules qui seront responsables de la dureté du tégument des Arthropodes.
b. Les proteines tannées
Elles représentent 25 à 75% du poids sec de la cuticule. Elles peuvent être révélées par les techniques de révélations de protéines comme le Western Blot. Afin de s’insérer dans la chitine qui est une molécule hydrophile, les protéines doivent présenter une portion hydrophile, comme l’arthropodine, et une portion hydrophobe, comme la sclérotine responsable de la dureté des téguments.
Arthropodine et sclérotine ne représentent pas des protéines particulières mais des groupes de protéines caractérisés par leurs propriétés assez identiques (poids moléculaire, charge des molécules qui provoque une mobilité plus ou moins importante en électrophorèse, solubilité, etc.) d’association à la chitine.
En réalité, arthropodines et sclérotines ne se différencient que par leur état physique, d’ailleurs leur séquençage en acides aminés ne révèle aucune différence. Les sclérotines sont des protéines cuticulaires banales, elles sont stabilisées par un tannage quinonique agissant sur les NH2 libres des protéines dans l’exocuticule, ceci n’affecte pas les arthropodines mais rend l’exocuticule plus dure.
c. Minéralisation
Chez les Crustacés et de nombreux Myriapodes, la dureté de la cuticule est fonction de la sclérotinisation des protéines cuticulaires comme chez les Insectes mais également d’une minéralisation par des sels de calcium. Ceci est exceptionnel chez les Insectes et n’a été signalé que chez quelques larves aquatiques.
On peut donc classifier les arthropodes en deux groupes :
- Les Arthropodes à squelette organique comme les Insectes, les Aranéides ou les Scorpionides
- Les Arthropodes à squelette calcifié comme les Myriapodes et les Crustacés
d. Les cuticulines
L’épicuticule est dépourvue de chitine, elle contient par contre de nombreuses substances assez mal connues. La cuticuline présente dans l’assise profonde résiste à l’action des solvants organiques et aux acides minéraux. C’est une lipoprotéine tannée par des quinones.
La couche cireuse est riche en un mélange de paraffines en C27 et C31, d’esters d’alcool supérieurs et d’acides gras non saturés en C26 et C30. Les chaines hydrophobes sont orientées perpendiculairement à la surface. Les pôles hydrophiles sont vers l’intérieur. Les pôles hydrophobes confèrent l’imperméabilité du tégument. Mettre un Insecte dans un détergeant provoque sa mort immédiate. Ceci est d’ailleurs utilisé dans de nombreux insecticides.
4. Conséquence de la présence de la cuticule
La cuticule forme un véritable exosquelette. De ce fait, il faut que l’animal s’adapte morphologiquement et physiologiquement.
Il existe une diversité des formations tégumentaires qui va avec la diversité morphologique des Arthropodes. Cependant, l’anatomie de ces animaux reste tout de même plus ou moins identique en fonction des classes.
Il n’existe aucun cil vibratil à la surface de tous les épidermes ectodermiques, notamment sur l’épiderme. Cependant, des formations ciliaires peuvent tout de même apparaitre de manière transitoire au cours de l’organogenèse des glandes ou des structures sensorielles.
Les mouvements relatifs des métamères ne sont possibles que grâce aux membranes articulaires où la cuticule est amincie et dépourvue d’exocuticule. L’abdomen des Insectescomporte des déformations très fréquentes dues aux variations volumétriques (alimentations et maturation des œufs chez les femelles). Les pleurites non sclérotinisés constituent, avec les membranes articulaires adjacentes, des zones souples et extensibles.
Pour une articulation, tous les mouvements ne sont pas possibles. En effet, les articulations mettent en jeu 2 condyles au maximum, on parle d’articulation monocondylaire et d’articulation dicondylaire. Ces articulations sont disposées selon deux modalités.
Les articulations intrinsèques où le condyle et la cavité glénoïde sont différenciés au sein de la membrane articulaire. Dans les articulations extrinsèques, les formations complémentaires se situent à l’extérieur des membranes articulaires.
Même si les articulations ne favorisent qu’au maximum deux plans de l’espace, la combinaison de ces articulations fait que les Arthropodes peuvent réaliser des mouvements complexes dans tous les plans de l’espace, on parle de mouvement omnidirectionnel.
Les muscles ont une action puissante, la cuticule a elle seul ne pourrait pas supporter les contractions, c’est pourquoi les faisceaux musculaires s’insèrent au niveau d’invaginations tégumentaires qu’on nomme apodèmes. L’ancrage des fibres musculaires se fait par des invaginations de la membrane plasmique et sarcoplasmique. De plus, de multiples desmosomes et microtubules forment un cytosquelette permettant la cohésion de l’ensemble. A chaque mue, cette association doit être renouvelée.
5. Mue et déroulement de la mue
Une fiche est disponible ici : http://www.fichier-pdf.fr/2012/07/11/la-mue-des-arthropodes/la-mue-des-arthropodes.pdf
La présence d’un exosquelette rigide et par définition inextensible, confère à la croissance des Arthropodes un caractère discontinu.
A la sortie de l’œuf, l’animal se développe, seulement il ne pourra le faire qu’en changeant sa cuticule et en grandissant pendant ce changement de cuticule.
Il arrive un moment où la cuticule devient trop petite pour la croissance ou pour les changements, l’animal va donc muer, la mue est un phénomène permettant la croissance discontinue de l’animal, elle se fait en plusieurs étapes.
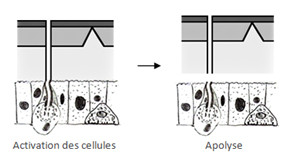
Tout d’abord il y a activation des cellules épidermiques, elles entrent en mitose, ceci est suivi d’un phénomène qu’on appelle apolyse, à ce moment la cuticule commence à se décrocher, ceci se fait plusieurs jours avant la mue.
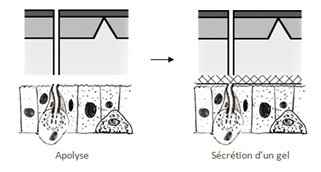
Ensuite il y a sécrétion par l’épiderme d’un gel de mue, ce gel est inerte car il est composé d’enzymes inactives. Cependant dès qu’il y a sécrétion de la nouvelle épicuticule par les cellules épidermiques, ce gel remplie d’enzymes va dégrader l’ancienne peau. Le gel va servir à recycler l’endocuticule, ces éléments vont être réintégrés dans les cellules pour fabriquer la nouvelle cuticule. Chez les crustacés les sources de calcium sont recyclées, il y a en effet des échanges d’ions permettant de réintégrer le calcium au niveau des cellules de l’épiderme, notamment l’échange de 2 K+ contre un ion Ca++ .

L’endocuticule ayant été dégradée va prendre l’aspect d’un liquide, qu’on nomme le liquide exuvial. Ce liquide sera réabsorbé en grande partie par l’épiderme. Entre temps, les oenocytes commencent à sécréter de la cire qui sera transmise aux cellules épithéliales, il y aura formation de canalicules des pores et dépôt de cires sur la nouvelle cuticule.
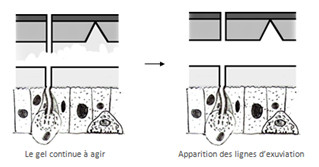
Le gel continuant à agir, Il y a des endroits où la cuticule devient très fine ces endroits sont appelés lignes d’exuviation, l’animal va se gonfler d’eau ou d’air et va casser la cuticule au niveau ces lignes, il y a alors exuviation.
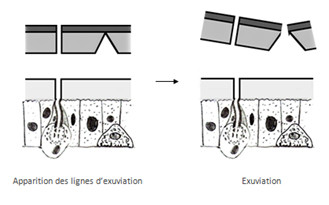
L’exuviation, encore appelée Ecdysis va permettre à l’animal de sortir de son ancienne cuticule. Pendant ce temps, les cellules épidermiques ont proliférées provoquant ainsi un plissement de l’épiderme. Après l’exuviation, l’épiderme va se déplier et l’animal va grandir, laissant derrière lui son ancien exosquelette qu’on nomme encore exuvie.
Le reste de la cuticule est alors synthétisé, il y a tannage des protéines, sclérotinisation et dépôt de minéraux. Les couches post ecdysiales sont après ça synthétisées. Pendant cette période, l’animal reste à jeun. En effet, il ne se nourrit qu’après que ça cuticule soit reformée, ce qui explique que la croissance en terme de poids a une allure assez différente à la croissance en longueur.
Le rapport des tailles successives prises par les Arthropodes au cours de leur développement post embryonnaire a en général à une valeur constante de 1,25. La longueur augmente donc du quart à chaque mue, c’est la Règle de Prizbram. Le poids quant à lui, double d’une mue à l’autre, c’est la règle de Dyar.
6. Les membranes articulaires
Les membranes articulaires se trouvent entre les différents métamères. Au niveau des soies et des pattes, la membrane articulaire possède une protéine particulière appelée la résine. De plus, elle ne possède pas d’exocuticule.
7. Les soies
La cuticule peut, chez certains animaux, être ornée de baguettes chitineuses qu’on nomme les soies, on en a deux types :
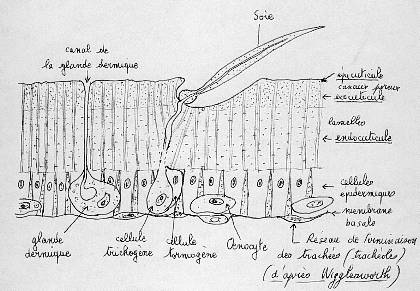
- Les soies pleines qui sont en général ornementales, cette soie est sécrétée par une cellule trichogène, cette cellule est entourée par une autre cellule qui la protège: la cellule tormogène. Les soies sont aplaties chez les Lépidoptères et ont un aspect écailleux. Chez l’abeille, les soies au niveau des pattes ont un rôle protecteur mais servent aussi à récolter le pollen.

- Les soies sensorielles qui ont la même organisation que les soies pleines mais sont reliées à une cellule nerveuse avec des dendrites qui sont dans la soie. La cellule nerveuse est entourée dans une cellule thécogène, le tout est entouré par la cellule trichogène puis par la cellule tormogène. Une soie ayant une cellule nerveuse portant plusieurs dendrites est appelée sensille. Les soies sensorielles peuvent être de deux types. Elles peuvent être gustatives d’une part et olfactives d’autre part.Les soies gustatives se trouvent souvent sur les pièces buccales et se terminent par un pore gustatif (non représenté sur le schéma) permettant à l’animal de gouter. Les soies olfactives s’ouvrent quant à elles par plusieurs pores (non représentés sur le schéma) qui permettent de capter une information olfactive, elles se trouvent au niveau des antennes.

B. Les appendices articulés
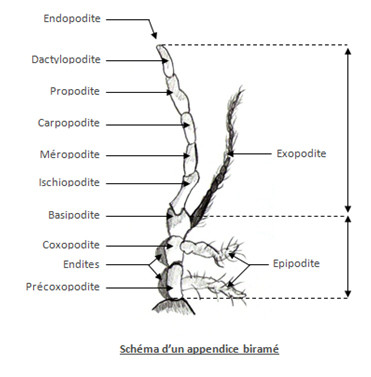
L’appendice des Arthropodes est soit uniramé soit biramé. Ceci fut un critère de classification, aujourd’hui ça ne l’est plus.
L’appendice est constitué de plusieurs parties, une base le protopodite qui articule l’appendice au corps et qui porte 2 rames chez les animaux possédant des appendices biramés, l’endopodite interne et l’exopodite externe. Les appendices uniramés ne possèdent pas d’exopodite. L’endopodite et l’exopodite, quand il existe, forment le télopodite.
Le protopodite est divisé en 3 articles, le précoxopodite, le coxopodite et le basipodite. Le précoxopodite, souvent soudé à la paroi du corps, et le coxopodite ont une partie interne, l’endite, et une partie externe : l’épipodite. La basipodite porte le télopodite.
Le télopodite est composé de deux rames distinctes quand la seconde existe, une rame interne ou endopodite comprenant 5 articles, l’ischiopodite, le méropodite, le carpopodite, le propodite et le dactylopodite. La seconde rame, ou exopodite, est externe et porte beaucoup plus d’articles.
Les appendices ont des formes et des fonctions très diverses. Organes sensoriels (antennes, pédipalpes ou antennules), pièces buccales ou pattes. Ils peuvent servir à la respiration, les épipodites portent alors des branchies, ou à la reproduction.
Autrefois, la classification officielle divisait les Arthropodes en uniramés et en biramés, les biramés étaient représenté par les crustacés. Seuls les crustacés possèdent donc un exopodite.
IV. Organisation et anatomie
A. Le tube digestif
1. La bouche entourée de pièces buccales
Chez les Chélicérates, il y a une paire de chélicères composée de 2 ou 3 articles en forme de pinces ou de crochets. Les chélicères sont souvent associées à des glandes à venin. La seconde paire d’appendices est la paire de pédipalpe composée de 6 articles qui ont un rôle sensoriel tactile.
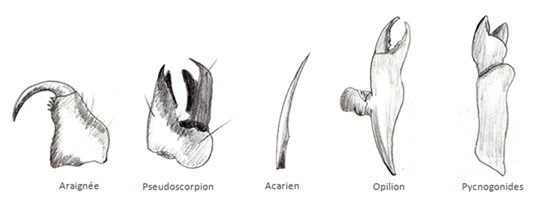
Chez les Mandibulates, les pièces buccales sont différentes en fonction des groupes.
a. Chez les Crustacés
Chez la langoustine, qui est un Crustacé, la tête porte 5 paires d’appendices. Les antennules (A1), les antennes (A2), les mandibules (Md), les maxillules (Mx1) et les maxilles (Mx2).
Les pièces buccales ne sont, par ailleurs, pas essentiellement portées par la tête, en effet elles sont composées d’une paire de mandibules (Md), une paire de maxillules (Mx1) et 3 paires de pattes mâchoires qui sont portées par le péréion (Pmx1, Pmx2 et Pmx3)
Schéma extrait d’une diapositive universitaire

b. Chez les Myriapodes
Les pièces buccales des Myriapodes sont portées par la tête, elles sont composées d’une paire de mandibules (Md), d’une paire de maxilles (Mx1 équivalentes aux maxillules des Crustacés) et d’une paire d’appendices qui fusionnent pour former le labium, ou lèvre inférieure (2Mx2 soudées équivalentes aux maxilles des Crustacés). En plus de ces pièces buccales, les myriapodes portent une paire de crochets associés à des glandes à venin, les maxillipèdes.

C. Chez les Insectes
Chez les Insectes, les pièces buccales sont portées par la tête, mais ne dérivent pas toute des appendices. En effet, on compte une lèvre supérieure, le labre, en forme de pelle qui dérive directement d’un métamère, d’ailleurs ce labre est surmonté par une membrane utile lors des déterminations, le clypeus. Les pièces buccales dérivant des appendices sont d’une part la paire de mandibule, qui est orientée vers l’intérieur, il y a les molaires qui broient et les incisives qui arrachent. D’autre part, on trouve une paire de maxilles, ou mâchoires antérieures qui portent chacune une palpe maxillaire composée de 5 articles. Enfin, les deux Mx2 fusionnent pour former le labium. En fonction des animaux les pièces buccales sont modifiées par le régime alimentaire.
La bouche d’un criquet est subdivisée en deux parties, l’hypopharynx, ou langue, délimite ces deux cavités. L’atrium buccal où s’ouvre le tube digestif et le salivarium où débouche les glandes salivaires.
Le criquet porte des pièces buccales de type broyeur.
Le type broyeur lécheur est surtout porté par les Hyménoptères comme les abeilles, Les paraglosses s’allongent pour former une sorte de paille qui permet aux insectes d’aspirer le nectar, les mandibules participant au type broyeur ne servent qu’à malaxer la cire et ne sont pas fortement développé.
Le type suceur est divisé en deux sous types :
- le type suceur maxillaire comme les papillons dont les mx1 vont réduire leurs galéas pour former une trompe.
- Le type suceur labial où le labium s’est allongé, il est retrouvé chez les mouches.
Le type piqueur suceur où les mandibules et les maxilles se transforment en stylet piqueur, ceci est bien visible chez les moustiques ou les Hémiptères.
2. Le tube digestif
Le tube digestif est limité par un épithélium simple et est composé de trois segments, un segment antérieur ou stomodeum, un segment médian ou mésentéron et un segment postérieur ou proctodeum.
- L’intestin antérieur : A l’entrée on a la bouche, on arrive dans le pharynx, puis l’œsophage, le jabot et le gésier, des coeca gastriques et des glandes externes lui sont associées. L’intestin antérieur est d’origine ectodermique, il est tapissé par une intima cuticulaires elle aussi renouvelée à chaque mue, il est ornementé de saillies qu’on appelle denticules ou râpes participant à la dégradation des aliments.
- L’intestin moyen: Il a un rôle dans la digestion et l’absorption. En effet, les cellules constitutives sécrètent des enzymes digestives et peuvent absorber les produits de la digestion. Il est d’origine endodermique. Parfois chez certains groupes, comme les Crustacés par exemple, le mésentéron est associé à une glande digestive particulière, l’hépatopancréas.
- L’intestin postérieur : Il se termine par le rectum et l’anus, d’origine ectodermique il possède aussi une intima cuticulaire. Il y a réabsorption d’eau et certains ions.
La paroi digestive est pourvue de muscles circulaires et longitudinaux qui assurent un mouvement du contenu du tube digestif vers l’extérieur, on parle de mouvement péristaltique.
A la limite entre le mésentéron et le proctodeum on a des tubes, les tubes de Malpighi, ce sont des organes excréteurs qui ne concernent que les insectes.
La morphologie et la physiologie du tube digestif est en relation avec le régime alimentaire de l’animal, il varie d’une espèce à l’autre, voire même à l’intérieur d’une même
espèce.

B. L’appareil respiratoire
La respiration chez les Arthropodes est complexe, ils colonisent tous les milieux on a donc plusieurs modes de respiration.
1. La respiration branchiale
Elle ne concerne que les Crustacés et les Mérostomes (limules) qui ont souvent des expansions foliacées au niveau du thorax. Ces branchies sont souvent reliées à la base des appendices, au niveau des épipodites, et servent à la respiration.
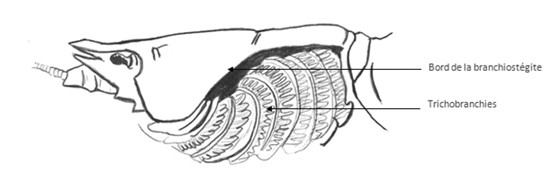
Chez les Crustacés supérieurs on a essentiellement deux types de branchies :
- les trichobranchies ou branchies filamenteuses : on a des filaments branchiaux en forme de tubes avec un vaisseau afférent et un vaisseau efférent. Les tubes sont séparés par un septum longitudinal, portant le nom de raphé, il va délimiter le trajet de l’hémolymphe. Les branchies sont formées par des cellules qu’on appelle les cellules piliers, ces cellules piliers forment des lacunes, l’hémolymphe circule dans chaque filament branchial afin d’arriver dans les lacunes, elle revient ensuite dans le vaisseau efférent. Un vaisseau a une paroi propre, une lacune n’en a pas, il s’agit d’un creux. Ce type de structure est retrouvé chez les langoustines ou les homards.
Fiche sur les branchies filamenteuses disponible ici : http://www.fichier-pdf.fr/2012/07/11/branchies-fillamenteuses/branchies-fillamenteuses.pdf
- Les phyllobranchies : ce sont des branchies aplaties en forme de lamelle, elles sont bordées par des canaux marginaux. Vue qu’elles sont très fines, ces branchies sont entourées par des cellules piliers et entre ces cellules se forment des lacunes, l’hémolymphe passera entre ces lacunes. Ce type de structure est retrouvé chez les crabes.
Fiche sur les branchies lamellaires disponible ici : http://www.fichier-pdf.fr/2012/07/11/branchies-lamellaires/branchies-lamellaires.pdf
Les branchies sont protégées par une cavité branchiale délimitée par un prolongement des tergites thoraciques, ceci est appelé le branchiostégite.
2. La respiration tégumentaire
Chez les Crustacés de petite forme, on a une respiration tégumentaire, souvent ceci se fait sur des surfaces très amincies, ici ceci se fait au niveau des appendices.
3. La respiration aérienne
a. Les pseudobranchies
Les pseudotrachées sont présentes au niveau des Isopodes terrestres, ce sont les seuls Crustacés qui vivent sur terre. Au niveau de leurs appendices, ils ont de petites surfaces d’échanges en forme de lacune sur leurs exopodites au niveau des pléopodes. Les trachées s’ouvrent au niveau de la base de l’appendice à sa face interne.
Les Ligies vivent quasiment toujours immergées mais ces autres isopodes portent aussi des pseudotrachées.
b. Le poumon des Arachnides
Ce poumon est une invagination du tégument, il a une forme globuleuse. Dans sa partie dorsale il y a des lamelles, les replis lamelleux, ces replis vont communiqués ensemble dans un atrium. Cet atrium communique avec l’extérieur par un stigmate. Entre les lamelles, on a des petites cellules qui forment des lacunes, c’est au niveau de ces lacunes que l’hémolymphe circule. Chez une araignée, il y a deux poumons (sauf chez les Mygales qui en ont 4), elles ont aussi un système trachéen. Le poumon est situé dans l’opisthosome.
Fiche sur les poumons des Arachnides disponible ici : http://www.fichier-pdf.fr/2012/07/11/poumons-des-araignees/poumons-des-araignees.pdf
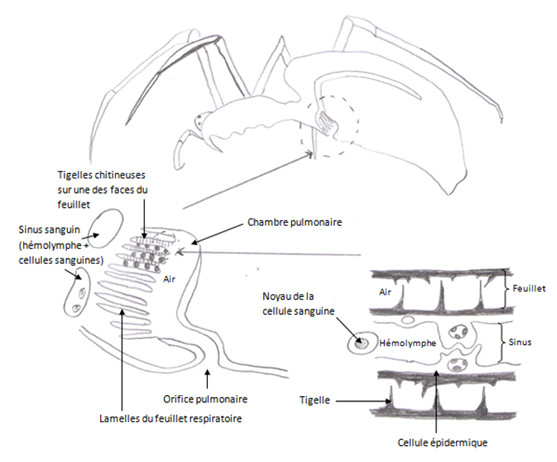
Les trachées des Araignées sont un peu spéciales car les trachéoles se terminent dans l’hémolymphe au lieu d’aboutir directement sur les cellules. Ici l’hémolymphe est un intermédiaire de transport local de l’oxygène.
c. Les trachées chez les Myriapodes et les Insectes
La respiration des Insectes est assurée par des trachées qui constituent un système très perfectionné. Ce sont des invaginations du tégument, cette fois ci de forme tubulaire, elles communiquent à l’extérieur par des stigmates. A l’intérieur de ces trachées, la cuticule forme à certains endroit des épaississements en spirale qu’on appelle ténidies, ceci empêche les tube de se déformer et donc de se fermer.
Fiche sur le système trachéen ici : http://www.fichier-pdf.fr/2012/07/11/le-systeme-tracheen/le-systeme-tracheen.pdf

Les trachées se ramifient en de nombreuses trachéoles, ces ramifications deviennent de plus en plus fines pour atteindre 3 à 5 µm en fin de ramification. Les dernières ramifications arrivent sur de grosses cellules, de forme étoilée, les cellules trachéolaires. De ces ramifications ce font des prolongements cytoplasmiques, les trachéoles. L’extrémité est aveugle et se termine directement dans les organes, l’oxygène arrive directement dans la cellule, les trachéoles repoussent le cytoplasme des cellules cibles, dans les trachéoles il y a un peu de liquide pour permettre la diffusion des gaz.
Chez les arthropodes les moins évoluées, chaque trachée provient d’un métamère, ces trachées sont assez peu ramifiées et sont indépendante les unes des autres. Par contre chez les arthropodes les plus évolués les trachées sont reliées les unes aux autres par des connexions transversales et longitudinales, ceci permet la circulation de l’air dans tout le corps.
Il existe 3 systèmes :
- Un système de ramification dorsale qui irrigue les muscles pariétaux et le cœur
- Un système latéral irrigant les viscères
- Un système ventral qui irrigue la chaine nerveuse et les muscles pariétaux
On trouve aussi des sacs aériens au niveau des muscles allaires chez les Ptérygotes, constituant des réserves d’air, ces sacs diminuent aussi la densité, la paroi est très mince, ce qui permet à l’animal par la contraction du corps de les remplir.
Plusieurs systèmes protègent le système respiratoire, l’obturation des stigmates par exemple. Ils s’ouvrent au niveau d’une chambre qu’on nomme atrium, cet atrium est bordé par un filtre et une valve, qui empêchent l’eau et les particules de rentrer dans les trachées. Ceci est une adaptation réussi chez les Arthropodes terrestres, car empêche les dessiccations et permet la respiration, comme il n’y a pas de transport de gaz, la diffusion est limitée par la longueur des trachées. Plus ces trachées sont longues, plus l’oxygène diffuse mal, ceci explique pourquoi les Arthropodes terrestres ont une taille limitée.
Lorsque l’Insecte mue, il renouvelle son appareil respiratoire.
d. La respiration des insectes aquatiques
Les larves utilisant l’O2 dissous dans l’eau
Les Ephémères ou les Odonates ont des larves qui vivent dans l’eau, en temps que larve ils n’auront pas de stigmate comme à l’état adulte, il y a un système particulier, le système trachéen existe déjà mais les stigmates sont clos. Par contre elles possèdent des trachéobranchies, l’oxygène y passe et sera transmis aux trachées. Ce sont des expansions tégumentaires au niveau de trachées localisées différemment en fonction des individus, ces expansions portent aussi des petites trachées, l’eau passe, l’O2 est capté au niveau de la trachéobranchie et est transmis aux trachées.
Fiche sur les trachéobranchies disponible ici : http://www.fichier-pdf.fr/2012/07/11/les-tracheobranchies/les-tracheobranchies.pdf
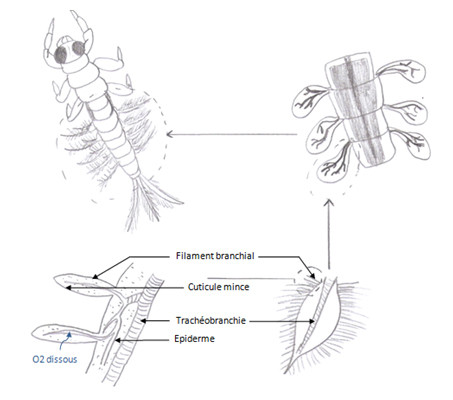
Les échanges peuvent se faire au niveau anal (Corbeilles branchiales). En effet un autre système est constitué par les branchies rectales. En coupe transversale de la larve, les branchies sont des invaginations de 6 replis au niveau du rectum. Par les contractions du cloaque, l’oxygène entre.
Fiche sur les corbeilles branchiales disponible ici : http://www.fichier-pdf.fr/2012/07/11/corbeille-branchiale/corbeille-branchiale.pdf

Les Insectes qui utilisent l’oxygène gazeux
Ces Insectes portent des systèmes particuliers, ils sont par contre tous aquatique. Tous les stigmates sont clos sauf la dernière paire abdominale. Ces stigmates s’ouvrent au niveau d’un siphon. Le siphon peut être lié à sa base ou à son extrémité au stigmate, il absorbe l’air et l’achemine vers les trachées.
Les larves ont un siphon respiratoire plus ou moins long avec ou non des crochets fixateurs à leurs bases, Les larves se tiennent alors verticalement ou obliquement sous la surface de l’eau. La disposition et la nature des soies, les proportions du siphon, la base du siphon sont utilisés en systématique.
Fiche sur la larve de Culex et son siphon disponible ici : http://www.fichier-pdf.fr/2012/07/11/larve-de-culex/larve-de-culex.pdf
La larve du Coléoptère Donacia pique le roseau à l’aide de l’éperon de son 8ème stigmate abdominal et prend l’oxygène dans les lacunes aérifères.
Certains autres insectes peuvent emmagasiner de l’air. Le dytique doit se débrouiller pour capter l’oxygène à la surface de l’eau. Pour rester plus longtemps sous l’eau, le dytique a trouvé une astuce. Il coince de l’air sous ses élytres. Quand la bulle d’air est vide, il remonte à la surface pour la recharger. Les notonectes portent quant à elles des soies hydrophobes qui coincent l’air dans leur dos, leur permettant de faire une réserve d’air.
e. Les Crustacés qui respirent hors de l’eau
Ces Crustacés portent une pièce buccale particulière, le scaphognatite, celui-ci bat normalement pour faire ressortir l’eau. Quand il n’y a plus d’eau elle bat dans l’autre sens, ce qui va oxygéner l’eau restante dans la cavité buccale et donc permet la respiration.
C. L’appareil circulatoire
L’appareil circulatoire n’est pas clôt, il est constitué d’un tube plus ou moins long constituant le cœur, ce tube porte des pores, les ostioles, il propulse l’hémolymphe d’arrière en avant, l’hémolymphe arrive dans les sinus et baigne les organes, le système qui ramène l’hémolymphe au cœur est très peu développé. En effet, il la ramène par des lacunes veineuses. Tout autour du cœur on a un sinus péricardique.
Chez quelques Crustacés et quelques larves d’Insectes on trouve de l’hémoglobine (notamment chez la larve de chironome) et de l’hémocyanine la couleur de l’hémolymphe n’a de rapport qu’avec ce que l’insecte mange. L’hémolymphe n’a aucun rôle respiratoire, elle ne porte donc pas de pigments colorés.
D. L’appareil excréteur
Excréter sert à la fois à maintenir l’équilibre des concentrations en soluté mais aussi au maintien dans l’organisme d’un volume d’eau idéal. Cela sert aussi à l’élimination des déchets du métabolisme et à l’élimination des substances étrangères ou de leurs produits.
Les systèmes excréteurs sont très variés selon les groupes, cependant ils utilisent l’ultrafiltration et le transport actif. L’ultrafiltration utilise les gradients de pressions, la pression entraine le passage du liquide à travers une membrane partiellement perméable, l’eau et les petites molécules solubles passent mais pas les grosses. Le transport actif consiste en un mouvement de soluté contre le gradient électrochimique grâce à un mécanisme nécessitant une dépense énergétique.
Le cœlome est réduit à certains organes excréteurs et les cavités gonadiques, les cœlomes n’ont plus de disposition métamérique, les néphridies n’en ont donc pas non plus. Chez les Arthropodes, l’appareil excréteur porte le nom de coelomoducte. Ces organes mettent en relation une cavité cœlomique avec l’extérieur. Ces organes sont retrouvés chez les Arthropodes marins, les Arthropodes terrestres ont des tubes de Malpighi.
1. Les glandes coxales
Les organes excréteurs sont appelés glandes coxales, elles sont situées à la base des pattes, on les trouve aussi au niveau des glandes antennaires ou mandibulaire, on a aussi les reins céphaliques des Myriapodes et de certains Insectes.
Fiche sur les glandes coxales disponible ici : http://www.fichier-pdf.fr/2012/07/11/glande-verte-de-l-ecrevisse/glande-verte-de-l-ecrevisse.pdf

Toutes ces glandes excrétrices ont un fonctionnement identique. Elles sont constituées de deux parties :
- Le saccule terminal d’origine cœlomique possédant des parois contenant des podocytes qui permettent l’ultrafiltration.
- Le coelomoducte qui est un tube contourné, qui peut se dilater en une vessie. Il débouche à la base de l’appendice qui lui a donné naissance (coxa, antenne …).
2. Les tubes de Malpighi
Les tubes de Malpighi sont présents chez les Insectes, les Myriapodes et les Arachnides. Ce sont des tubes grêles qui baignent dans l’hémolymphe, ils débouchent dans le tube digestif entre le mésentéron et le proctodeum, l’extrémité distale est aveugle, leur nombre varie en fonction des espèces, de 2 à 2500. Ils forment de l’urine pâteuse, elle est fabriquée par sécrétion au niveau de la partie distale et réabsorption au niveau proximal. Il n’y a donc pas d’ultrafiltration, cette urine est évacuée par l’anus.
3. Les néphrocytes
Ces cellules sont présentes chez tous les Arthropodes terrestres, elles peuvent être individuelles ou en amas et sont dispersées dans tout le corps. Chez les crustacés elles sont disposées dans l’hémocoele le long de l’axe branchial. Chez les Insectes on les trouve au niveau du sinus péricardique. Ces cellules sont là pour dégrader les substances complexes qui ne peuvent pas être dégradées dans tes tubes de Malpighi. Les néphrocytes absorbent les produits et les dégradent, ces nouveaux produits sont largués dans l’hémolymphe et réabsorbés par les tubes de Malpighi.
4. Les reins d’accumulation
Ce sont des structures où sont stockés les métabolites azotés à vie chez les Arthropodes terrestres. Par exemple, il y a un endroit chez le criquet qui garde ces métabolites, le tissu adipeux, c’est le siège de l’accumulation de bases puriques. Les tubes de Malpighi peuvent, chez certaines espèces, servir au stockage définitif. Les néphrocytes sont aussi capable de stocker des déchets azotés.
E. L’appareil genital
D’une façon générale, les Arthropodes sont des animaux à sexes séparés, ce sont des animaux gonochoriques, certains sexes sont morphologiquement très différents. L’échange de gamète se fait par accouplement. L’accouplement est très varié en fonction des groupes et même des espèces. Les gonades sont associées à des organes annexes, il y a aussi des organes servant à la reproduction ou au stockage des spermatozoïdes.
1. Chez les Crustacés
L’appareil génital est situé dans le péréion, les gonades sont paires et soudées sur la ligne médiane, et prennent souvent l’aspect d’un Y, les gonades sont dans une cavité représentée par un résidu de cœlome.
Chez le mâle les canaux déférents s’ouvrent à la base des coxopodites des Pe8 (péréiopodes 8). Pour la femelle, les oviductes s’ouvrent à la base des coxopodites des Pe6.
Les spermatozoïdes, quand ils sont émis sont regroupés en spermatophores, ils sont captés par les gonadopodes, les pl1 et 2 (pléopodes 1 et 2) et ils sont déposés sur les sternites thoraciques à la base des pe6 là où sortent les ovocytes. Il y a fécondation.
Les œufs sont pondus et fécondés dès la sortie des oviductes. Soit ils sont libérés dans l’eau de mer, soit ils sont collés au niveau d’un substrat, soit les œufs sont accrochés au pléopodes 3 à 5 par des sécrétions d’une glande, les glandes cémentaires.
Les œufs des Crustacés sont riches en vitellus, la segmentation est donc difficile, elle est donc partielle. L’œuf libère une larve Nauplius.
2. Chez les insectes
Les sexes sont séparés et le dimorphisme sexuel est plus ou moins accentué. D’une manière générale les appareils génitaux sont battis sur la même structure.
Les gonades et les gonoductes sont pairs, ces structures sont d’origine mésodermique. Elles hébergent les cellules germinales dont la finalité est d’aboutir aux gamètes.
Le conduit génital est impair et médian, il est d’origine ectodermique et est aussi commun.
Chez les Insectes on a un appareil copulateur externe, le génitalia. Chez le mâle, il sert au rapprochement des sexes et au transfert des spermatozoïdes, les pièces sont très différentes et permettent de ne pas avoir de fécondation entre deux espèces.
L’appareil femelle est composé de 3 valves et est appelé ovipositeur.
La gonade mâle est composée de tubes séminifères, ils sont reliés par un pédicelle à un tube commun: le spermiducte. Parfois, les testicules sont accolées pour former une masse testiculaire, la partie basale du spermiducte peut être dilatée en vésicule séminale, ils se terminent par le canal éjaculateur, en commun avec l’appareil copulateur (pénis ou aedegus). On a des glandes accessoires qui permettent d’agglomérer les spermatozoïdes en spermatophores et de faire du liquide séminal. L’aedegus est souvent associé à deux formations chitineuses, les épiphalus.
La gonade femelle est composée de 2 ovaires, faite par un certain nombre d’ovarioles (ou ovigères). A l’intérieur, on a des gamètes dont la taille est de plus en plus grande. Ça débouche sur un pédicelle qui peut être fermé par un bouchon épithélial qui est détruit à chaque ovulation. Chaque ovariole est composé de 4 parties :
- Un filament terminal fait d’un prolongement de l’ovariole, ils servent à garder la position de l’ovariole, accroché à la partie dorsale. L’ensemble de ces filaments est appelé ligament suspenseur.
- Le germarium, qui contient des cellules appartenant a deux séries distinctes, les cellules germinales primordiales dont l’évolution donne les ovocytes et les cellules préfoliculaires d’origine mésodermique pour former un follicule, ces dernières sont appelées aussi cyctocytes.
- le vitellarium, il s’agit de la zone de croissance de l’ovocyte, on trouvera une série d’ovocyte linéaire dont la taille sera croissante.
- Le pédicelle avec le bouchon épithélial
Les deux oviductes latéraux vont fusionner en un vagin médian, les œufs en fonction des espèces, qui sont pondus seuls ou en amas, descendent et sont fécondés dans la chambre génitale par les spermatozoïdes stockés dans la spermathèque.
Les œufs sont imbibés d’une substance sécrétée par les cellules cholérétiques. Ils sont collés à un substrat, il existe un appareil de ponte et l’oviscapte pour capter les spermatozoïdes.
L’œuf est centrolécithe et la segmentation est superficielle, l’œuf libère une larve.
3. Chez les arachnides
Les sexes sont séparés et à l’exception des Acariens, l’orifice génital est situé à la face ventrale du 2ème somite abdominal (8ème somite du corps).
4. Chez les mérostomes
Les sexes sont séparés mais l’appareil reproducteur a la même organisation selon que l’on se trouve chez le mâle ou la femelle. La gonade est unique, elle est située au dessus du tube digestif dans la moitié postérieure du prosoma et dans l’abdomen. Elle est constituée par un réseau symétrique de tubules ovariens ou spermatiques. Les gamètes sont conduits par un court gonoducte vers le gonopore.
5. Chez les Pycnogonides
Les Pantapodes sont aussi gonochoriques, la présence d’une structure particulière permet de distinguer le mâle de la femelle, en effet la femelle porte des ovigères.
La gonade est unique, chez les mâles et les femelles, elle se trouve dorsalement au tube digestif, elle a la forme d’un U dirigé vers l’arrière. Les branches de cette gonade envoient des prolongements dans chaque appendice. Les orifices sexuels se trouvent à la base des Coxae. A maturité, les œufs migrent dans les fémurs des gonopodes (pattes portant les gonopores).
F. Le développement post embryonnaire
1. Chez les Crustacés
La croissance se fait de façon discontinue, elle est entrecoupée de mues, chez les Crustacés la croissance est indéfinie, elle continue toute la vie sauf chez une espèce l’araignée de mer maya qui s’arrête de grandir après une mue de puberté.
Il existe deux sortes de développement :
a. Le développement condensé
Le jeune a directement la forme de l’adulte.
b. Le développement dilaté
La vie larvaire comprend un grand nombre de stade morphologiquement distinct, la forme adulte apparait progressivement. Plusieurs stades larvaires (5) sont nécessaires à la mise en place de la forme adulte :
- le premier est le nauplius. Entre chaque stade, on a un certain nombre de mue qui conduisent à changer progressivement l’animal entre la larve nauplius et la larve métanauplius.
Ce premier stade est caractéristique de la majorité des Crustacés marins, il s’agit de la forme larvaire la plus primitive. Il s’agit d’une forme planctonique de 0,5 mm de long qui ne présente aucune trace de segmentation, son corps est ovoïde et présente deux soies caudales. Cette forme porte cependant 3 paires d’appendices dont les deux postérieures sont biramées, ils correspondent d’avant en arrière aux antennules, aux antennes et aux mandibules. Les deux appendices biramés (antennes et mandibules) présentent beaucoups de soies. Le tube digestif se situe à l’arrière de deux masses de mésoderme. Il commence au niveau de la bouche qui se situe entre les deux antennes. Elle présente une lèvre supérieure et une lèvre inférieure, respectivement appelées épistome et métastome. Le tube digestif se termine par l’anus qui s’ouvre entre les deux soies caudales.
L’appareil circulatoire est pour le moment absent. Cependant, il existe un appareil excréteur constitué de deux glandes coxales à la base des antennes, on parle aussi de glandes antennaires.
A la face dorsale, il y a présence d’un œil en position médiane et antérieure, il est constitué de trois ocelles. Le système nerveux est quant à lui rudimentaire, il présente un amas ganglionnaire supra-œsophagien (futur protocérébron) et 3 masses ganglionnaires sous-œsophagiennes qui sont en fait les neuromères correspondant aux trois paires d’appendices.
Après études embryologique, on sait que le nauplius est représenté par la fusion de l’acron de 4 somites et du telson.
- Le métanauplius suit après plusieurs mues le stade nauplius (1 jour et demi entre le stade nauplius et métanauplius). Il mesure 0,4 à 0,6 mm. Son corps est en forme de raquette, il possède 4 ébauches de métamères postmandibulaires dont le bourgeon des maxillules, des maxilles et des deux premiers péréiopodes. Le telson se termine par une structure particulière qu’on nomme furca de deux expansions latérales. L’organisation interne se perfectionne pendant environ le temps de 7 mues.
- La larve protozoé est le troisième stade et apparait moins de 3 jours après l’éclosion. A ce stade apparaissent deux régions distinctes : une région antérieure de forme ovalaire et recouverte d’une carapace, ainsi qu’une région postérieure de forme cylindrique et se terminant par une furca très bien marquée. Les appendices maxillules, maxilles, pmx1 (péréiopodes 1) et Pmx2 (péréiopodes 2), acquièrent leur forme définitive. La larve mue pendant 4 à 5 jours et atteint une taille de 1,5 mm. Pendant ces mues, les yeux continuent à se différencier et on voit apparaitre la segmentation de la région postérieure
- La larve zoé apparait 2 à 3 jours plus tard, cet état larvaire est caractérisé par la présence des péréiopodes 3 (Pmx3) qui est biramé et par l’apparition des 5 derniers segments thoraciques. L’abdomen ne porte qu’une paire d’appendice au niveau du dernier somite, ces appendices fusionneront avec la furca du telson pour former une nageoire caudale.
- La larve mysis fait suite au stade zoé. Après 3 jours, elle acquit les pléopodes, les segments thoraciques sont recouvert d’une carapace et sont incorporés au céphalothorax.
2. Chez les insectes
La croissance est discontinue et limitée, la dernière mue est la mue imaginale, car elle libère la dernière forme de l’Insecte qui est appelé imago.
- Chez les Amétaboles
Les Amétaboles sont les Insectes qui ne présentent pas de métamorphose, Le jeune ressemble donc à l’adulte, il y aura croissance de l’antenne et développement de ses différents segments. Lors de la mue imaginal l’animal acquière des gonades matures. Il existe chez ces Insectes des mues post imaginal qui vont alterner les cycles de reproduction.
- les Hétérométaboles
Les Hétérométaboles regroupent les Insectes qui ont des métamorphoses complètes et incomplètes. On parle respectivement des Holométaboles et des Hémimétaboles. Il existe aussi un autre groupe, les Paurométaboles qui n’ont pas réellement de métamorphose mais juste les ailes qui se développent. L’œuf libère une larve différente de l’adulte, les gonades n’apparaissent qu’à la mue imaginale.
- Les Paurométaboles, la larve est identique à l’adulte mais il va y avoir développement des ailes qui va être progressif, l’ébauche de l’aile est visible dés la naissance, on parle d’Exoptérygotes.
- Les Hémimétaboles sont aussi des Exoptérygotes, l’ébauche de l’aile est visible, mais il y a des différences entre l’adulte et la larve, autant morphologique que comportementale. La mue imaginale entraine des changements très importants en rapport avec le mode de vie (aquatique à terrestre dans la plupart des cas), les ébauches allaires ne se développent qu’à la mue imaginale.
- Les Holométaboles sont Endoptérygotes, les ailes n’apparaissent qu’au dernier stade de la vie, le stade nymphal. A la fin de ce stade nymphal, il y a la mue imaginale qui est la véritable métamorphose.
3. Chez les arachnides
Le développement se fait sans métamorphose. Le cœlome se segmente en deux bandes de mésoderme qui se segmentent en autant de cavités que de métamères externes. Ceci les fait se rapprocher des Annélides.
a. Cas des Scorpionides
Les scorpions sont vivipares ou ovovivipares. Les ovovivipares possèdent des œufs dont le développement se fait dans la lumière des tubules ovariens. Les espèces vivipares ont des œufs dépourvus de vitellus, ces œufs se développent isolément dans des diverticules ovariens en relation avec un caecum digestif maternel. Il s’agit d’un équivalent au cordon ombilical chez les Mammifères.
Le développement est extrêmement lent (quelques mois voire une année) et aboutit à la naissance de 6 à 19 jeunes qui grimpent immédiatement sur le dos de leur mère pour accomplir leur L1 (premier stade larvaire qui dure une semaine). Les jeunes quittent ensuite progressivement la mère pour devenir indépendant, en un an et un nombre de mue variable selon les espèces, ils deviennent adultes.
b. Cas des Aranéides
La femelle pond jusqu’à 3 000 œufs répartis en un nombre variable de cocons soyeux qui peuvent être, selon les espèces, transportés par la mère comme Pisaura mirabilis ou Pardosa amentata, ou fixés sur différents substrats. Chez L’épeire diadème Araneus diadematus, le cocon est fixé au dessus de la toile, à l’abri des prédateurs, il en est de même pour les espèces du genre Tegenaria, à ceci près que ces espèces forment des toiles en forme de tube, les cocons sont ainsi protégés à la fois par le « terrier » mais aussi par la mère qui garde l’entrée.
Les œufs présentent un vitellus centré, on parle d’œuf centrolécithes, le développement commence par une segmentation partielle. Après la sortie de l’œuf, les jeunes font leur première mue et sortent du cocon. La dispersion des petits se fait souvent par le vent, le jeune est alors accroché à un long fil de soie, on parle du fil de la Vierge.
Le nombre de mue pour arriver au stade est variable selon l’espèce et la taille. Seules les femelles des Mygalomorphes muent après avoir atteint le stade adulte.
G. Système nerveux et organes des sens
1. Le cerveau
Le cerveau des Arthropodes résulte de la fusion de l’Archicérébron aussi appelé protocérébron et des deux premiers neuromères, on les appelle deutocérébron et tricérébron. Le protocérébron est en relation avec les organes visuels (yeux composés, ommatidies ou ocelles), il contient donc de gros ganglions optiques. Le corps central, composé du pont protocérébral et des corps pédonculés, forme des centres d’associations et d’intégration, ceux-ci sont très développés chez les Insectes sociaux.
Le Deutérocérébron est en relation avec les antennules et les antennes, il contient les ganglions antennaires ou antennulaires. Ces centres olfactifs symétriques sont réunis au dessus de l’œsophage, par une structure qu’on appelle commissure supra-œsophagienne.
Le tricérébron est encore sous l’œsophage, la céphalisation de ce neuromère est incomplète. Il innerve les antennes, les chélicères ou le labre selon la classe d’Arthropodes.
L’organisation de ce cerveau est très proche de celui des annélides. D’ailleurs, embryologiquement, l’origine est la même, il s’agit de structures préorales sauf dans le cas du tricérébron qui est postoral et qui devient préoral au cours du développement de certains Arthropodes. En effet, chez certains, notamment les Crustacés Branchiopodes, les Collemboles ou les Hémiptères, la céphalisation du tricérébron n’est pas achevée et il occupe toujours une position postorale.
Le tricérébron est en relation avec le système sympathique antérieur aussi appelé stomogastrique. Ce système innerve le stomodeum (partie antérieure du tube digestif).
2. La chaine nerveuse
La chaine nerveuse est ventrale, comme chez tous les hyponeuriens, elle comporte une paire de ganglions par métamère qui sont associés entre eux par une commissure transversale et avec la paire antérieure et postérieure par des connections longitudinales. Cependant via la tagmatisation, les ganglions d’une même paire sont soudés entre eux et les connections longitudinales associées aussi. Chaque ganglion est à l’origine de 3 paires de nerfs.
Les ganglions portent beaucoup de cellules neurosécrétrices, les produits de sécrétions seront directement déversés dans l’hémolymphe au niveau des organes neuro-hémaux. Ces substances contrôlent divers processus comme par exemple la mue, la digestion, l’excrétion, ou la vitellogenèse.
3. Les organes des sens
On distingue deux types d’organes sensoriels selon qu’ils donnent des informations internes ou externes à l’organisme, on parle respectivement d’organes proprioceptifs ou intéroceptifs et d’organes extéroceptifs.
Dans les organes extéroceptifs on peut distinguer des organes photorécepteurs, gustatifs, olfactifs, auditifs ou thermosensibles. Ces éléments sont toujours construits sur la même base de 3 éléments :
- Des cellules sensorielles, des cellules épidermiques sont reliées par des neurofibres sensorielles à un centre nerveux, constituent alors le début d’un arc réflexe (neurone afférent, interneurone et neurone efférent).
- Une formation cuticulaire dont la structure est variable en fonction de la propriété de l’organe, son but est de transmettre l’information du milieu extérieur à la cellule sensorielle. Précédemment, nous avons décrit la structure d’une soie sensorielle, le principe reste identique, la soie capte l’information et la fournit au neurone.
- Des éléments annexes, une ou plusieurs cellules épithéliales ont un rôle dans la sécrétion de cuticule ou dans la protection de la cellule nerveuse, c’est typiquement le cas de la cellule trichogène qui entoure et protège la terminaison dendritique de la cellule nerveuse au niveau de la soie.
L’ensemble de ces constituants forment ce qu’on appelle une sensille. Ce sont des structures d’origine ectodermique, elles sont donc toutes périphériques. Ce sont elles qui vont conditionner les actions de l’animal.
Les organes photosensibles ou ommatidies fonctionnent aussi sur le même principe. On a des cellules épidermiques spécialisées, les cellules rétiniennes disposées en une seule assise et dont le prolongement forme le nerf optique. Cette assise est superposée d’une formation cuticulaire qui joue le rôle de lentille dont le rôle est de concentrer la lumière. On distingue selon le nombre de formations cuticulaires, les ocelles qui ne possède qu’un dioptre pour l’ensemble des cellules sensorielles et les yeux composés qui ont autant de formations cuticulaires que de cellules sensorielles.
Bibliographie
DEMANGE, Jean-Marie. Myriapodes [En ligne]. In : Encyclopaedia Universalis. Paris : Encyclopaedia Universalis S.A, 2008. Disponible sur http://www.universalis-edu.com/ (consulté le 25 janvier 2012)
FOREST, Jacques. Crustacés [En ligne]. In : Encyclopaedia Universalis. Paris : Encyclopaedia Universalis S.A, 2008. Disponible sur http://www.universalis-edu.com/ (consulté le 25 janvier 2012)
IAVCHA. Le tégument [en ligne]. Disponible sur http://www.iavcha.ac.ma/benazoun/entomologie-generale/docs/tegument.html (consulté le 21 janvier 2012)
LEGENDRE, Roland, VACHON, Max. Arachnides [En ligne]. In : Encyclopaedia Universalis. Paris : Encyclopaedia Universalis S.A, 2008. Disponible sur http://www.universalis-edu.com/ (consulté le 25 janvier 2012)
LEGENDRE, Roland, VACHON, Max. Arthropodes [En ligne]. In : Encyclopaedia Universalis. Paris : Encyclopaedia Universalis S.A, 2008. Disponible sur http://www.universalis-edu.com/ (consulté le 19 janvier 2012)
LEGENDRE, Roland, VACHON, Max. Chélicérates [En ligne]. In : Encyclopaedia Universalis. Paris : Encyclopaedia Universalis S.A, 2008. Disponible sur http://www.universalis-edu.com/ (consulté le 19 janvier 2012)
“The Tree of Life Web Project”. Disponible sur http://tolweb.org/tree/phylogeny.html (consulté le 21 janvier 2012)
WIKIPEDIA. Chelicerata [en ligne]. In : Wikipedia l’encyclopédie libre. Etats Unis : Wikimedia Foudation, Inc. Mise à jour le 28 septembre 2011. Disponible sur http://fr.wikipedia.org/wiki/Chelicerata (consulté le 21 janvier 2012)
WIKIPEDIA. Pycnogonida [en ligne]. In : Wikipedia l’encyclopédie libre. Etats Unis : Wikimedia Foudation, Inc. Mise à jour le 28 mai 2011. Disponible sur http://fr.wikipedia.org/wiki/Pycnogonida (consulté le 21 janvier 2012)
WIKIPEDIA. Lame basale [en ligne]. In : Wikipedia l’encyclopédie libre. Etats Unis : Wikimedia Foudation, Inc. Mise à jour le 28 novembre 2011. Disponible sur http://fr.wikipedia.org/wiki/Lame_basale (consulté le 21 janvier 2012)
BEAUMONT, André, CASSIER, Pierre. Biologie animale : Des protozoaires aux métazoaires épithélioneuriens. 3ème édition, Paris : 2000, Dunod. ISBN : 2 10 048660 8
Crédits photos et illustrations
Illustrations et photos
http://www-museum.unl.edu/research/entomology/workers/graphics/Latreille.JPG : Latreille
{kind=link}
http://sciencextra.files.wordpress.com/2008/11/limuleplage.jpg : regroupement de limule
{kind=link}
http://clione.ru/wp-content/gallery/underwater/pavuk.jpg : Pantopode
{kind=link}
http://i34.servimg.com/u/f34/11/13/26/35/reptil12.jpg : Scolopendre
{kind=link}
© Alexander Semenov : http://clione.ru/
Illustrations utilisées pour la confection des schémas
http://locust.cirad.fr/images_locusts/mpat136.jpg Téguments des arthropodes
{kind=link}
http://lemondedesphasmes.free.fr/IMG/cache-450x378/mue-fig3-et-4-450x378.jpg Téguments des arthropodes
{kind=link}
http://aramel.free.fr/Sensillecuticule.jpg Téguments des arthropodes
{kind=link}
http://s3.e-monsite.com/2010/09/28/09/resize_550_550//a2.png Téguments des arthropodes
{kind=link}
http://s3.e-monsite.com/2010/09/28/09/resize_550_550//a1.png Schéma d’une cellule épidermique
{kind=link}
http://www.astrosurf.com/luxorion/Bio/cellule-animale.jpg Schéma d’une cellule épidermique http://micro.magnet.fsu.edu/cells/nucleus/images/nucleolusfigure1.jpg Schéma d’une cellule épidermique
{kind=link}
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Gastrulation.png/300px-Gastrulation.png embryologie
{kind=link}
http://www.infovisual.info/03/img_fr/067%20Oeuf%20f%E9cond%E9.jpg embryologie
{kind=link}
http://cours.cegep-st-jerome.qc.ca/101-nya-m.b/Classification/coelome_2.JPG embryologie
{kind=link}
http://s1.e-monsite.com/2010/07/13/11/resize_550_550//ARTHRO7.jpg Métamère d’un arthropode
{kind=link}
Métamère, © 2005 Encyclopaedia Universalis France S.A
La plupart des illustrations ont pour base des schémas tirés de ce livre : BEAUMONT, André, CASSIER, Pierre. Biologie animale : Des protozoaires aux métazoaires épithélioneuriens. 3ème édition, Paris : 2000, Dunod. ISBN : 2 10 048660 8